草地早熟禾氮代谢及与根茎扩展的相关性研究
时间:2023-02-15 16:00:09 来源:千叶帆 本文已影响人
袁娅娟,白小明,朱雅楠,张毓婧,闫玉邦,张才忠,李玉杰
(甘肃农业大学草业学院,草业生态系统教育部重点实验室,甘肃省草业工程实验室,中⁃美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
草坪是城市绿化中种植面积最大的地被植物,作为多年生禾本科植物,草地早熟禾(Poa pratensis)因其绿期长、再生能力和耐修剪性强的特性,成为人工草坪的重要组成成分[1]。作为典型的根茎型禾草,通过根茎不断产生新的分蘖株来进行种群扩充,是草地早熟禾再生能力强的关键所在[2-3]。氮在植物生命活动中占据首要地位,堪称生命元素,是叶绿体、核酸、蛋白质以及很多次生代谢产物的重要组成成分[4]。氮素对于植株的器官建成、生理代谢、干物质生产和种子产量的形成都起着重要作用[5-6]。因此,了解草地早熟禾氮代谢规律对草坪草的建植与养护管理具有十分重要的现实意义。
氮代谢是植物体内最基本的两大代谢过程之一,研究植物氮代谢产物及其相关酶活性在不同环境下的动态趋势,能在一定程度上反映植株对环境的响应与调节能力[7]。酶作为代谢网络运行的调控单元,对代谢控制至关重要。硝酸还原酶是氮素同化的第一个酶,也是限速酶,谷氨酰胺合成酶(GS)、谷氨酸合成酶(GOGAT)和谷氨酸脱氢酶(GDH)参与氮代谢调节,GS/GOGAT 循环实现了无机氮的有机化,它们共同构成氮代谢运转与循环[8]。Brain 等[9]认为氮素通过两条途径调节植物生长发育,一是调节植物体内营养水平进而影响发育,二是通过一系列信号物质调节植物代谢。已有研究证实,小麦植株体内氮代谢与分蘖发生、衰亡及其成穗密切相关[10]。杨文钰等[11]发现,营养代谢水平对分蘖成穗具有重要作用,小麦生育前期旺盛的碳代谢、生育后期旺盛的氮代谢均有利于分蘖的发生。关于草地早熟禾的相关研究表明,单一氮素条件供给下,适量施氮可提高草地早熟禾细胞抗氧化能力、草坪质量,并促进其生长[12-13];
水氮互作条件下,适量灌水和氮肥施用对推迟草地早熟禾枯黄和提前返青有显著作用[14]。目前,对于氮素的研究热点集中在氮素添加对植物的影响[15-16]以及土壤氮储量上[17-18],对于小麦和水稻的氮素动态变化规律也有研究,但时期大多集中在花后或籽粒灌浆期[19,21],有关草地早熟禾返青后各生育时期内氮代谢酶活性的研究报道较少。为此本研究以根茎扩展能力不同的3 个野生草地早熟禾为材料,在大田栽培条件下,通过对6 个连续生育时期的根茎扩展及氮代谢指标的测定,探究含氮化合物与根茎扩展的相关性,以期为草地早熟禾根茎扩展调控机理及合理施肥提供理论依据。
1.1 试验地概况
试验地位于甘肃省兰州市甘肃农业大学草坪实训基地内,E 103°3′、N 36°48′,属于中温带气候区,平均海拔1 517 m,年平均气温10 ℃,年均降水量362 mm,年蒸发量1 623 mm。土壤以壤土为主,土层深厚。土壤基础理化性状:有机质2.34%,速效氮40.37 mg/kg,速 效 磷45.62 mg/kg,速 效 钾75.3 mg/kg,pH 值为7.5。
1.2 试验材料
试验材料为3 个根茎扩展能力不同的野生草地早熟禾,采自甘肃境内(表1)。

表1 供试3 个野生草地早熟禾Table 1 Materials information of 3 testedPoa pratensis
1.3 试验设计
试验采取大田种植方式,播前施磷酸二铵30 g/m2作为底肥,于2019年6月12日种植于甘肃农业大学草坪实训基地。采用株行距70 cm×70 cm 进行穴播,每穴约50 粒种子。出苗两周后第1 次间苗,3 周后第二次间苗,5 周后选取生长健壮,均匀一致,无病虫害的植株定苗。以试验地内50%植株外部形态发生显著变化为标准,分别于种植第2年的返青期、拔节期、抽穗期、开花期、成熟期和枯黄期进行6 次取样,取样时间9∶00~11∶00。其中每个材料取8 株,用蒸馏水洗净并吸干水分,4 株用于根茎扩展指标测定;
其余4 株全部取一级根茎,将其称重、锡箔纸包裹,液氮迅速冷冻后置于-80 ℃超低温冰箱保存,用于相关生理指标测定。
1.4 指标测定与方法
1.4.1 分蘖数 测定草地早熟禾地面枝条的数量,用蘖/株表示。
1.4.2 根茎数 测定草地早熟禾地下根茎数,用个/株表示。
1.4.3 地上和根茎生物量 采用烘干称重法,将地上与地下根茎分开,洗净,置于105 ℃杀青30 min,70 ℃烘干至恒重,电子天平称其干重,用g/株表示。
1.4.4 硝态氮含量、游离氨基酸含量和硝酸还原酶活性 参照《植物生理学实验指导》测定[22]。
1.4.5 谷氨酰胺合成酶(GS)、谷氨酸合酶、谷氨酸脱氢酶活性 参照王小纯等[23]的方法测定,以单位时间内每mg 样品540 nm 处的吸光值表示GS 活性,单位为A/(g·h);
以单位时间内每g 样品催化NADH 的微摩尔数表示GOGAT和GDH活性,单位为μmol/(g·min)。
1.4.6 生物量增长率 用绝对增长率(AGR)表示,计算公式如下[24]:

式中:Wi、Wi+1分别表示ti、ti+1时刻的生物量。
1.5 数据分析
采用SPSS 26.0 软件,用Duncan 法进行方差分析和差异显著性检验,Excel 2019 绘制图表,试验结果为3 次重复的平均值。
2.1 草地早熟禾根茎扩展指标动态变化
2.1.1 分蘖数与根茎数动态变化 随生育时期推进,3 个材料分蘖数变化规律一致,均呈逐渐增加趋势(图1⁃A)。返青 期分蘖数 最少,LZYZ、DXWY 和DXAD 分 别 为361.00、290.00、217.00 蘖/株,LZYZ比DXWY 和DXAD 高22.48%和66.36%;
3 个材料分蘖数在枯黄期均达最大值,分别为1 196.00、827.33、440.00 蘖/株,LZYZ 比DXWY 和DXAD 高44.56%和171.82%。返青期至抽穗期是草地早熟禾分蘖数快速增长时期,抽穗期3 个材料分别较返青期增加了176.08%、127.13%和65.90%;
开花期后草地早熟生长速率减缓,至枯黄期3 个材料仅增加4.73%、9.80%和7.98%。
根茎数变化规律与分蘖数一致,随着生育时期推进均呈渐增趋势(图1⁃B)。在返青期根茎数最少,LZYZ、DXWY 和DXAD 分别为115.33、75.67、45.33个/株,LZYZ 比DXWY 和DXAD 高52.42% 和154.41%;
返青期后持续上升直至枯黄期达整个生育时期最大值,此时3 个材料分别为1 131.00、626.33、347.00 个/株,LZYZ 比DXWY 和DXAD 高80.57%和225.94%。返青期至抽穗期是根茎数快速增加时期,相比于返青期,抽穗期3 个材料分别增加662.14%、544.94%和429.41%;
开花期后增长速率逐渐下降,至枯黄期3 个材料仅增加15.88%、20.91%和20.91%。方差分析显示,整个生育时期内,分蘖数LZYZ 与DXAD 差异均不显著,根茎数除返青期、拨节期、抽穗期LZYZ 与DXAY 差异显著外,其余均差异不显著。
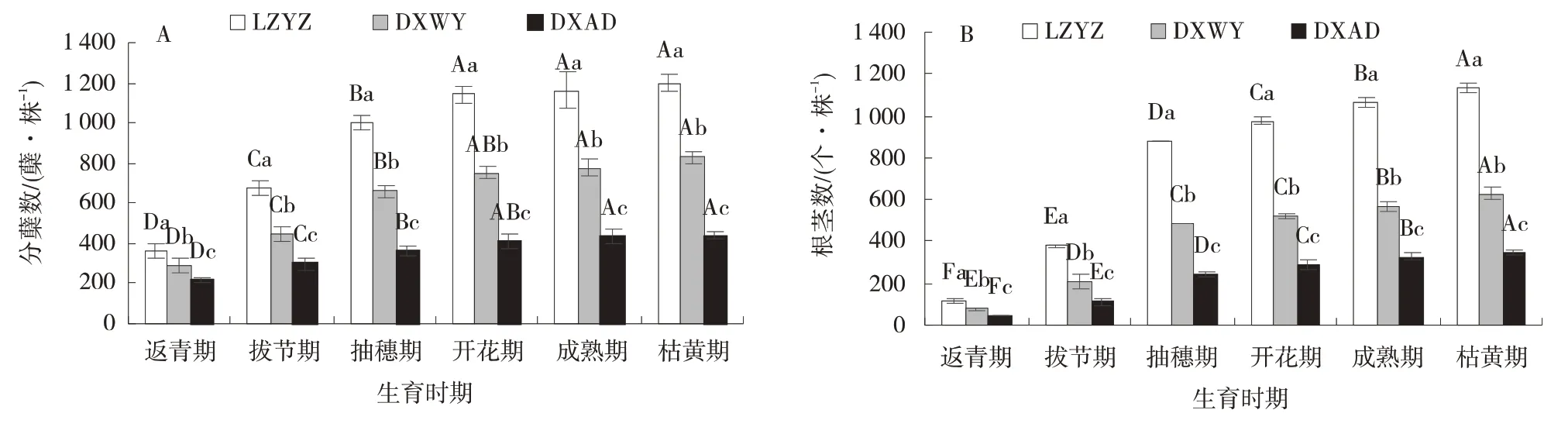
图1 不同生育时期分蘖数与根茎数Fig.1 Dynamic changes of tiller number and rhizome number
2.1.2 地上生物量动态变化 供试草地早熟禾地上生物量随生育时期推进变化趋势一致,均呈单峰曲线变化,自返青期后持续上升,在成熟期达到峰值,之后迅速下降(图2⁃A)。返青期,3 个材料地上生物量 均 处 于 最 小 值,LZYZ、DXWY 和DXAD 分 别 为21.94、20.49、8.59 g/株,LZYZ 比DXWY 和DXAD高7.08%和155.55%;
成熟期,3 个材料地上生物量分 别 为 232.23、162.80、136.26 g/株,LZYZ 比DXWY 高42.65%,比DXAD 高70.43%;
成熟期后3 个材料地上生物量迅速下降,分别减少24.23%、36.02% 和56.23%;
枯 黄 期LZYZ 比DXWY 高68.94%,比DXAD 高195.04%。方差分析显示,LZYZ 与DXWY 整个生育时期中除成熟期差异不显著外(P>0.05),其他时期差异显著(P<0.05),LZ YZ 与DXWY 除开花期和柘黄期差异显著外,其他时期差异不显著。且均表现为LZYZ>DXWY>DXAD。
草地早熟禾地上生物量绝对增长率随生育时期推进呈单峰曲线变化,成熟期前为正值,成熟期后为负值(图2⁃B)。峰值出现在拔节期至抽穗期,先于地上生物量达到最大值,3 个材料分别为136.22、78.71、68.33 g/(株·d),LZYZ 明显高于DXWY 和DXAD。抽穗期后逐渐减小,于成熟期后出现负值,3 个材料分别为-56.27、-58.64、-76.62 g/(株·d)。

图2 不同生育时期地上生物量及绝对增长率Fig.2 Dynamic changes of aboveground biomass and absolute growth rate
2.1.3 根茎生物量动态变化 供试草地早熟禾根茎生物量随生育时期推进与地上生物量的变化有所不同,但3 个材料间变化趋势一致,均呈“升⁃降⁃升”的“N”字型曲线(图3⁃A)。返青期根茎生物量最低,LZYZ、DXWY 和DXAD 分 别 为2.35、1.68、0.80 g/株;
随生育时期推进持续上升至开花期达第1 个峰值,此 时LZYZ 比DXWY 高75.82%,比DXAD 高423.89%;
开花期至成熟期3 个材料均下降,LZYZ 和DXAD 分别从33.42 g/株下降至29.67 g/株,6.38 g/株下降至4.54 g/株,两个生育时期间均差异不显著(P>0.05),DXWY 从19.01 g/株下降至8.94 g/株,差异显著(P<0.05);
在枯黄期3 个材料根茎生物量迅速增加,达整个生育时期最大值,LZYZ、DXWY 和DXAD 分别为75.06、44.04、20.80 g/株。方差分析显示,整个生育时期LZYZ 均显著高于DXWY 和DXAD,3 个材料根茎生物量一直表现为LZYZ>DXWY>DXAD。
根茎生物量绝对增长率在生长季的中前期和末期为正值,而生长季的后期为负值(图3⁃B)。根茎生物量AGR 有两个较大值,分别为拔节期至抽穗期和成熟期至枯黄期,3 个材料分别为26.33、13.48、4.42 g/(株·d)、45.39、35.11、16.26 g/(株·d);
负值为开花期至成熟期,3 个材料分别为-3.75、-10.07、-1.84 g/(株·d)。
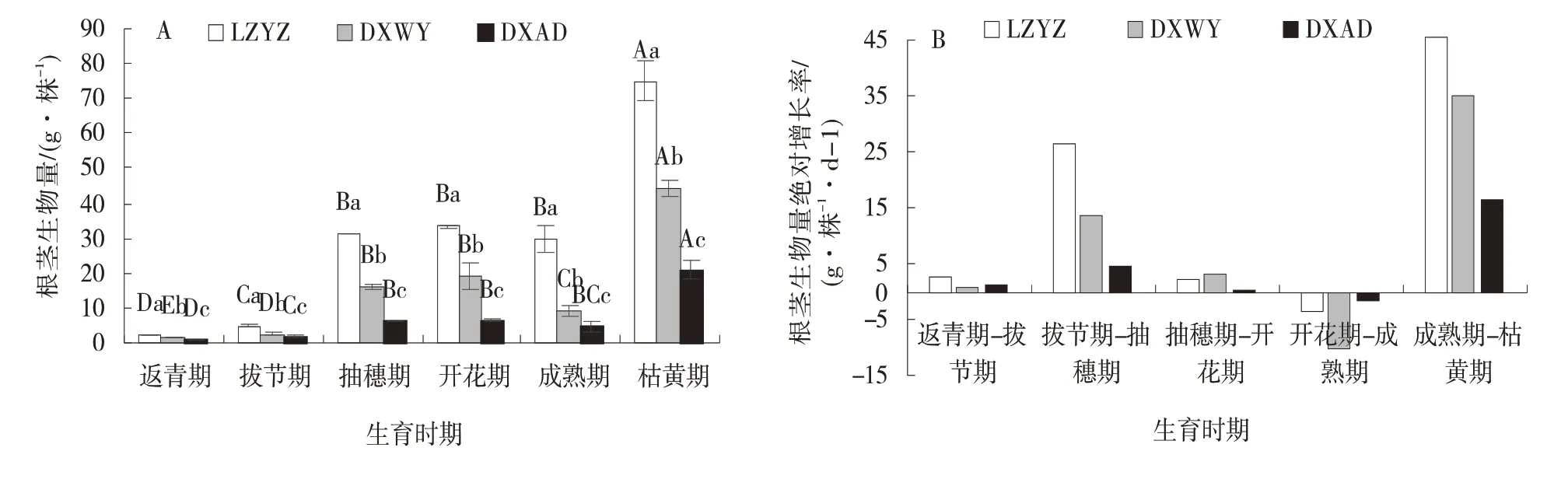
图3 不同生育时期根茎生物量及绝对增长率Fig.3 Dynamic changes of rhizome biomass and absolute growth rate
2.2 含氮化合物动态变化
2.2.1 硝态氮含量变化 随生育时期推进,3 个草地早熟禾材料NO3-⁃N 含量整体上变化规律一致,均呈单峰曲线变化(图4)。返青期3 个材料NO3-⁃N 含量均 处 于 较 低 水 平,LZYZ、DXWY 和DXAD 分 别 为350.20、250.34、240.58 μg/g,LZYZ 比DXWY 高39.89%,比DXAD 高45.56%。拔节期迅速上升,达到整个生育时期最大值,DXWY 增长最快,相较于返青期增加63.11%;
DXAD 增加48.40%;
LZYZ 增加幅度最小,仅为30.46%;
此时期3 者NO3-⁃N 含量差异不显著(P>0.05)。拔节期后3 个材料均持续下降,在枯黄期为整个生育时期最小值,分别为307.53、217.31、186.24 μg/g,三者差异显著(P<0.05)。N 含量变化趋势相似,均呈单峰曲线,只是峰值略微滞后于NO3-⁃N,出现在抽穗期(图6)。返青期3 个材料NR 活性均为整个生育时期最小值,LZYZ、DXWY 和DXAD 分别为10.28、8.90、5.69 μg/(g·h),三者差异显著(P<0.05);
返青期后均持续上升至抽穗期达到峰值,3 个材料分别增加了65.56%、77.07% 和161.13%,LZYZ 比DXWY 高8.07%,比DXAD 高14.69%,3 个材料差异不显著(P>0.05);
峰值过后整体呈下降趋势,直至枯黄期。
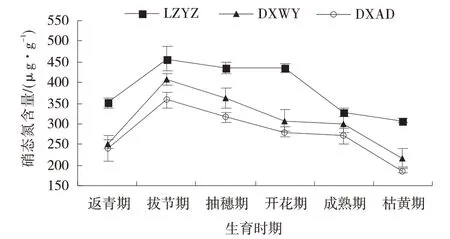
图4 不同生育时期草地早熟禾的硝态氮含量Fig.4 Changes of nitrate nitrogen content content inPoa pratensis
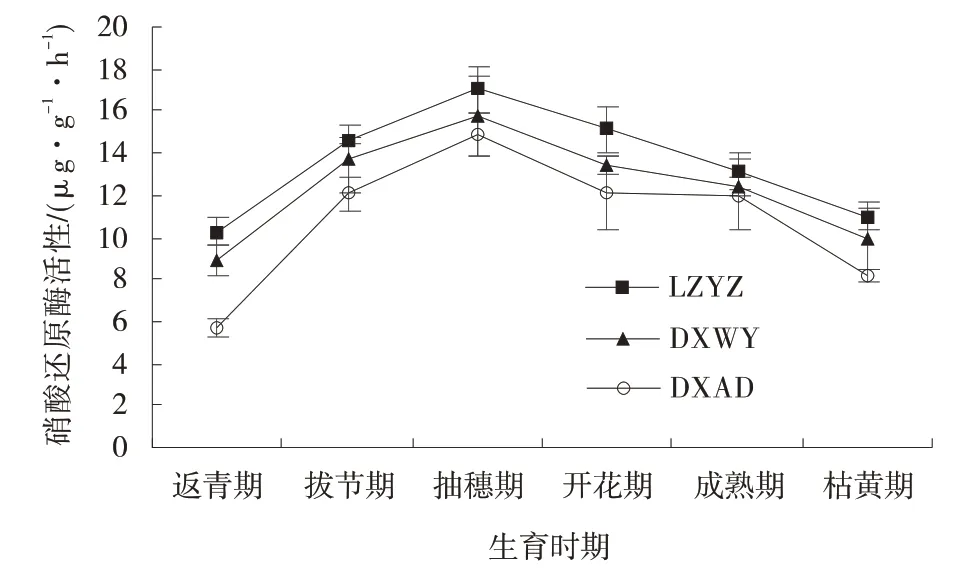
图6 不同生育时期草地早熟禾硝酸还原酶活性Fig.6 Changes of nitrate reductase activity inPoa pratensis
2.2 游离氨基酸含量变化
游离氨基酸含量随生育时期推进均呈“升⁃降⁃升”的变化规律(图5)。返青期3 个材料均处于整个生育时 期 最 小 值,LZYZ、DXWY 和DXAD 含 量 分 别 为18.65、19.65、17.52 μg/g,三 者 差 异 不 显 著(P>0.05);
返青期后持续上升至抽穗期出现整个生育时期最大值,三者游离氨基酸含量分别增加371.37%、303.77% 和 240.15%,此 时 LZYZ 比 DXWY 高10.77%,比DXAD 高13.99%,LZYZ 显 著 大 于DXWY 和DXAD(P<0.05);
抽穗期后迅速下降直至成熟期下降趋势停止。在枯黄期三者含量均有所上升,但增加幅度不同,相比于成熟期,LZYZ 增加65.50%,DXWY 增加28.36%,DXAD 增加56.39%,此时游离氨基酸含量表现为LZYZ>DXWY>DXAD,三者差异显著(P<0.05)。

图5 不同生育时期草地早熟禾游离氨基酸含量Fig.5 Changes of free amino acid content inPoa pratensis
2.3 氮代谢相关酶活性变化
2.3.1 硝酸还原酶活性动态变化 NR 活性与NO3-⁃
2.3.2 GS 活性变化 随生育时期推进,草地早熟禾GS 活性变化呈双峰曲线,峰值出现抽穗期和成熟期(图7)。返青期3 个材料GS 活性均处于整个生育时期 最 低 值,LZYZ、DXWY 和DXAD 分 别 为10.85、7.10、4.51 A/(g·h),持续上升至抽穗期达到第一个峰值,此时3 个材料酶活性分别为30.92、25.87、20.82 A/(g·h),LZYZ 比DXWY 高24.32%,比DXAD 高48.47%。开花期均有不同程度下降,在成熟期达到第二个峰值,LZYZ 上升幅度最大,相比于开花期增加19.72%;
DXAD 增加12.39%,DXWY 增加最少,仅为2.61%;
LZYZ 显著大于DXWY 和DXAD(P<0.05)。枯黄期均迅速下降,DXWY 与DXAD 之间差异不显著(P>0.05)。

图7 不同生育时期草地早熟禾GS 活性Fig.7 Changes of GS activity ofPoa pratensis
2.3.3 GOGAT 活性 GOGAT 活性与GS 活性变化趋势相似,均呈“升⁃降⁃升⁃降”的“M”型曲线变化,峰值出现在抽穗期和成熟期(图8)。返青期LZYZ、DXWY 和DXAD 酶 活 性 分 别 为2.27、1.73、1.70 μmol/(g·min),三者差异不显著(P>0.05);
返青期至拔节期酶活性缓慢增长,两个时期间差异不显著(P>0.05);
拔节期后迅速上升至抽穗期达到第1 个峰值,也是整个生育时期最大值,此时3 个材料分别为10.04、7.37、5.02 μmol/(g·min),LZYZ 比DXWY 高36.17%,比DXAD 高100.17%,三者差异显著(P<0.05);
在开花期迅速下降至较低水平;
于成熟期出现第2 个峰值,此时3 个材料酶活性分别为7.78、5.20、4.30 μmol/(g·min),较 开 花 期 分 别 增 加136.00%、70.49% 和80.04%,均显著小于第1 个峰值(P<0.05),且三个材料间差异显著(P<0.05),在枯黄期酶活性均下降且材料间差异不显著(P>0.05)。

图8 不同生育时期草地早熟禾GOGAT 活性Fig.8 Changes of GOGAT activity ofPoa pratensis
2.3.4 GDH 活性变化 整个生育时期中,供试3 个材料GDH 活性变化趋势一致,只是材料间活性高低略有差异(图9)。3 个材料在返青期酶活性较低,LZYZ、DXWY 和DXAD 分 别 为0.50、0.23、0.26 μmol/(g·min),返青期至抽穗期3 个材料酶活性基本维持在这一水平,且前期材料间表现为LZYZ 大于DXWY 和DXAD。抽穗期至成熟期酶活性迅速上升,3 个材料分别提高558.61%、1 527.78%、1413.04%,LZYZ 和DXWY 均显著高于DXAD(P<0.05);
成熟期至枯黄期GDH 活性略有下降,但两个生育时期间差异不显著(P>0.05)。

图9 不同生育时期草地早熟禾GDH 活性Fig.9 Changes of GDH activity ofPoa pratensis
2.4 草地早熟禾根茎扩展各指标增长速率与含氮化合物的相关性
整体上草地早熟禾根茎扩展能力与硝态氮和游离氨基酸含量、NR、GS 和GOGAT 活性正相关,与GDH 活性负相关(表2)。其中分蘖数增长率与硝态氮显著正相关,相关系数为0.496。根茎数增长率与硝态氮、游离氨基酸、NR 和GOGAT 极显著正相关(P<0.01),相 关 系 数 分 别 为0.677、0.663、0.658 和0.714;
与GS 活性显著正相关(P<0.05),相关系数为0.485。地上生物量增长率与硝态氮、NR 和GOGAT极显著正相关(P<0.01),相关系数为0.648、0.703 和0.602;
与游离氨基酸和GS 显著正相关(P<0.05),相关系数分别为0.528 和0.537。GDH 活性与分蘖数和地上生物量增长率呈极显著和显著负相关,相关系数分别为0.711 和0.582。根茎生物量增长率与以上指标相关性均不显著(P>0.05)。

表2 根茎扩展指标增长率与含氮化合物的相关系数Table 2 Correlation coefficient between growth rate of rhizome index and nitrogen compound
无性系的生长伴随着构件数量的增加及所占空间的增大。本研究中草地早熟禾分蘖数和根茎数在整个生育时期均呈渐增趋势,但返青期至抽穗期增长最快,抽穗期后增长速率减缓,这与孙建华等[25]发现大部分草地早熟禾在4、5月出现分蘖高峰期,极少数能延续至6月中旬的结果一致;
也与其地上部及根部最适生长温度为15~20 ℃和10~18.3 ℃的特点相吻合[26],表明草地早熟禾无性系的扩张活动在开花期前已基本完成。开花期是植物由营养生长转为生殖生长的标志,开花后花、穗和种子等生殖器官成为植物的生长及营养物质的输入中心,营养生长基本停止[27]。生物量随生育时期的动态变化具有明显趋势,许志信等[28]发现早熟禾地上生物量随其生育时期推进呈增加趋势,峰值出现在9月初;
郑伟等[29]表明,在牧草整个生长季,地上生物量变化呈单峰型,峰值出现在8月上旬。大量研究表明,地下生物量随生育时期或季节推进主要变化趋势有两种:单峰型和双峰型曲线变化[30]。本试验中,草地早熟禾地上生物量呈单峰曲线变化,峰值出现在成熟期,根茎生物量呈“N”字型变化,与高寒草甸地下生物量变化趋势相同[30]。
NO3-⁃N 是植物体内重要的营养元素,根系是吸收硝态氮的主要器官。郭彦青[31]对杨树的研究发现,全年中,杨树根部NO3--N 含量呈单峰曲线变化,在4月13日达到峰值后持续下降。本研究中,NO3--N含量变化与上述研究一致,呈单峰曲线,峰值出现在拔节期,拔节期后由于生长中心的转移及后期根系活力下降等原因,含量持续下降至与返青期相近水平。游离氨基酸既是氮的同化产物又是蛋白质分解的产物,是源库间实现氮素分配、转移、再分配的主要形式,可以反映植物体内参与转移的氮素基本情况[32]。王宇通[33]研究发现,在全叶马兰一个生长周期中,根部游离氨基酸呈钟型变化,早期和末期含量较少,中期含量高。本试验中,草地早熟禾前期变化与全叶马兰相似,但在枯黄期表现为上升趋势,这一现象与多年生禾本科植物奇岗[34]的趋势一致。由于植物在生长的不同时期追求对氮素利用最大化,而叶片中的氮素含量往往最高,当秋冬叶片衰老、枯萎凋落时,叶绿体解体,叶绿素分解释放出大量氮素,实现氮素向其他器官的转移,从而防止由叶片脱落导致的氮素流失,称为养分回流,转移的器官因植物而异。品种间氮代谢酶活性的差异是品种对于氮素吸收、利用差异的 基础[19],本研究中,LZYZ 的NR、GS、GOGAT 和GDH 酶活性在整个生育时期均高于DXAD,表明与其余两个材料相比,LZYZ 有着更强的氮素同化能力。
氮素对植物生长尤为重要,尤其是对分蘖的发生起决定性作用,人们通常通过增施氮肥来提高作物分蘖能力[35]。田广丽等[35]发现,水稻的相对分蘖速率与氮浓度呈极显著正相关。刘杨等[36]发现,低浓度氮水平下,分蘖芽处于休眠状态;
而较高浓度的氮可以解除分蘖芽休眠,促进其生长。相关研究表明,氮素含量增加一方面使水稻分蘖中玉米素与生长素含量增加,脱落酸含量降低,从而促使分蘖萌发;
另一方面,植物体内较高的氮素含量显著提高了叶中的光合速率,促进植物体内非结构性碳水化合物的积累,为分蘖生长提供足够同化物,因此植物体内氮素通过调节激素和干物质生产两方面来调控分蘖的发生[37]。Eg⁃gens 等[38]研究发现,高比率硝态氮可以促进早熟禾分蘖及增加地上部分干重;
张鹤山等[39]研究表明,氮肥施量与草坪草地上生物量、草坪密度和草坪高度显著正相关。本试验中相关性分析显示,分蘖数、根茎数及地上生物量增长率均与硝态氮和游离氨基酸极显著或显著正相关,与上述结论一致。
返青期至抽穗期是草地早熟禾根茎扩展的主要阶段;
拔节期至抽穗期是地上生物量快速积累积累阶段;
根茎生物量有两个快速积累阶段,分别为拔节期至抽穗期和成熟期至枯黄期。草地早熟禾生长发育过程中有两个氮代谢活跃的时期,分别为抽穗期和成熟期,相比于DXWY 和DXAD,LZYZ 有更强的氮素同化能力。草地早熟禾根茎扩展能力受含氮化合物及相关酶活性的影响,根茎中NO3--N 和游离氨基酸含量越多,NR、GS 和GOGAT 活性越高,GDH 活性越低,越有利于其根茎扩展。






