大别山区两种柿资源种实表型性状的多样性
时间:2023-02-18 11:40:08 来源:千叶帆 本文已影响人
杨 旭,徐 阳,龚榜初*,谢其均
(1.中国林业科学研究院亚热带林业研究所,浙江 杭州 311400;
2.河南省国有商城黄柏山林场,河南 商城 465350)
植物表型性状具有观察直观、调查简便、直接有效等优点,通过对表型变异程度和变化规律的研究,可以揭示植物对不同环境的适应能力,因此,在遗传变异、遗传多样性、良种选育等研究中被广泛应用[1]。在所有的表型性状中,叶片等营养器官受外界环境影响较大,表型变异的稳定性相对较低;
而作为生殖器官的果实和种子的表型变异则具有较高的稳定性,是植物种群长期遗传和环境自然选择的综合体现[2]。对柿属(Diospyros Linn.)柿(D.kaki Thunb.)[3]、浙江柿(D.glaucifolia Metc.)[4]、金枣柿(D.sp.)[5]、罗田甜柿(D.kaki Thunb.)[6]等植物果实和种子表型性状已有众多研究。植物的表型分化来源于生存环境的时空异质性、物种长期适应进化所累积的遗传物质及群体内个体、器官对生境的响应差异[7]。由于植物自身对所处生境的耐受性存在差异,使得一些植物分布范围广泛,另一部分只能分布在特定的区域内[8]。局域种和广布种在生物学和生态学特征方面存在着差异[9],而目前对同域分布的柿属不同植物表型性状的变异规律仍然缺乏了解。
罗田甜柿为我国特有的原产甜柿资源,仅分布于大别山区[10],是由单宁含量少、酒精脱氢酶较多的野生柿演变后,经过自然淘汰形成,至今已有900余年的栽培历史[11]。罗田甜柿是与日本甜柿独立起源、相对独立分化的2个群体,具有完全不同的自然脱涩特点和遗传背景,作为育种亲本可明显缩短育种进程,提高完全甜柿的育种效率[10]。许多学者关注这种特有的甜柿资源,分别以罗田甜柿为父、母本,创制出具有中国甜柿自然脱涩特点的完全甜柿新种质[12-13];
还通过胚乳培养、秋水仙碱处理等方法获得十二倍体植株[14-15]。野柿(D.kaki Thunb.var.silvestris Makino)为柿的变种,广布于我国中部和云南、广东、广西北部及江西、福建等省区,生于山地自然林或次生林中,或山坡灌丛中,垂直分布约达1 600 m[16]。野柿实生苗可作栽培柿树的砧木,用野柿为砧木嫁接‘太秋’甜柿的果实平均单果质量和果实内在品质显著高于其它砧木上的果实[17]。野柿是大别山区分布最广、数量最丰富的柿属植物资源。野柿与罗田甜柿在大别山区的分布区存在一定的重叠。本文旨在对大别山区罗田甜柿和野柿资源较全面收集的基础上,对其果实和种子的表型性状进行测定分析,揭示2种柿资源变异的丰富程度及变异规律,为该地区罗田甜柿和野柿资源的保存利用及构建核心种质提供科学依据。
1.1 试验材料
于2020年10月调查大别山区的罗田甜柿和野柿资源。采集地点包括安徽省霍山县但家庙镇、大别山国家地质公园白马尖园区,安徽省金寨县燕子河镇、天堂寨镇,河南省商城县黄柏山森林公园,湖北省罗田县惠兰山、骆驼坳镇、云架山、平湖乡、河铺镇等乡镇及麻城县盐田河镇等地区。采集地的地理位置为 115°15′~116°26′ E,30°2′~31°28′ N,海拔43~806 m,采集点基本囊括罗田甜柿的主要分布区。
为尽量减少频繁种质交流带来的影响,采集的罗田甜柿植株均为树龄100 a以上的大树。为保证试材具有足够的代表性,各采集点选取植株尽量在30株以上,植株间隔30 m以上,不足30株的则全部采集。每个植株采集成熟度基本一致的果实30个以上,于采集当天空运回实验室进行样品处理及各项指标的测定。
1.2 测定方法
采集的罗田甜柿和野柿果实主要形态见图1。以《柿种质资源描述规范和数据标准》[18]为参考依据,对采集的柿果实进行形态学测定。用游标卡尺测量果实横径、果实纵径、柿蒂大小、萼片大小、种子长和种子宽,用电子天平测量单果质量和种子质量,数出每个果实内种子数,并计算果形指数和种形指数;
选取成熟度基本一致的果实,用手持糖量计测量每个果实的可溶性固形物含量,用手持式硬度计测量果实的硬度[18]。

图1 罗田甜柿 (左)和野柿(右)果实形态Fig.1 Fruit phonotype of D.kaki (left)and D.kaki var.silvestris(right)
1.3 数据处理
以不同群体中柿种质资源的13个种实表型性状作为特征量,采用SPSS19.0软件中的Oneway Anova程序进行分析,分别计算不同性状的平均值、平方和、均方、标准差、标准误差、F值和显著水平,并统计频数及分组频率,进行正态分布型拟合。正态分布拟合分析中的偏度和峰度(削度)估计,由该软件提供的程序计算得到[19]。
对不同群体罗田甜柿和野柿的表型数据进行巢式方差分析,线性模型为:

式中:Yijk为第 i个群体第 j个家系第 k个观测值,μ 为总平均值,Si为群体效应(固定),T(i)j为群体内家系效应(随机),ɛ(ij)k为试验误差[20]。
计算群体间和群体内的方差分量,进而计算出群体间的表型分化系数(VST):

采用shannon′s多样性指数(H′)评价罗田甜柿和野柿各表型性状的遗传多样性[21]。依据不同性状的平均值和标准差,将表型划分为10个等级。shannon′s多样性指数计算公式为:

式中:Pi为各表型性状第i级别内的表型占总数的比例。
2.1 罗田甜柿和野柿种实表型性状分布规律
正态分布的分布型拟合分析获得的峰度和偏度值绝对值越接近于0,说明拟合分布型越接近标准分布。对罗田甜柿和野柿不同群体果实和种子表型性状频数的正态分布型拟合结果(表1)可知:除罗田甜柿的果实硬度外,罗田甜柿和野柿其余性状频数分布与正态拟合非常贴近。从2种柿资源不同性状统计的峰度和偏度值看,2种种质群体的平均偏度和平均峰度绝对值都较小,说明群体的取样及表型性状观测值的测定符合随机交配大群体法则,具有较好的代表性。

表1 柿种质种实性状测定值正态分布型拟合的偏度和峰度统计Table 1 Frequency distribution of quantitative characters index of persimmon
2.2 罗田甜柿和野柿种实表型性状群体间和群体内变异特征
对罗田甜柿和野柿不同群体种实表型性状进行方差分析,结果(表2)表明:F值的大小在一定程度上表明表型性状在群体间和群体内差异程度的高低。罗田甜柿13个种实表型性状中除萼片大小、种子数、种形指数、可溶性固形物含量在群体间达到极显著水平,种形指数在群体内达到极显著水平,萼片大小在群体内达到显著水平外,其余性状在群体间和群体内均未达到显著水平。野柿群体表型性状中,果形指数、柿蒂大小、萼片大小、种子质量、果实硬度等指标在群体间达到极显著水平,果实纵径在群体间达到显著水平,另有果实硬度在群体内也达到极显著水平。
植物不同表型性状的极值往往反映该性状在不同环境条件下的进化及适应性潜力。罗田甜柿13个表型性状的极大值为极小值的 1.42(柿蒂大小)~4.07(单果质量)倍。依据极大值与极小值的差异倍数,各表型性状的进化及适应性潜力从高到低分别为:单果质量 >种子质量 > 种子数 > 种子宽 > 果实硬度 > 种子长 > 萼片大小 > 果实纵径 > 可溶性固形物含量 > 果实横径 > 种形指数 >果形指数> 柿蒂大小。野柿13个表型性状的极大值为极小值的1.48(果形指数)~5.54(种子质量)倍。各表型性状的进化及适应性潜力从高到低分别为:种子质量 > 种子数 > 单果质量 > 果实硬度 > 果实纵径 > 可溶性固形物含量 > 萼片大小 >果实横径 > 种子宽 > 种子长 > 柿蒂大小 > 种形指数>果形指数 。
变异系数的大小反应了表型性状的离散程度,变异系数越小,离散程度越小,性状越稳定。罗田甜柿不同群体间果实和种子的数量性状变异系数为6.41%~25.85%,平均变异系数为13.78%,其中,变异系数较大的是种子数、果实硬度及单果质量,变异系数较小的是柿蒂大小、果形指数、果实横径、种形指数及果实纵径。不同群体野柿果实和种子表型性状的变异系数为10.92%~38.36%,平均变异系数为22.14%,显著高于罗田甜柿,其变异系数大小为:果实硬度 >单果质量 > 种子数 >种子质量> 种子宽 > 萼片大小 >可溶性固形物含量 >种子长 > 果实纵径 > 果实横径 > 种形指数 >柿蒂大小 > 果形指数。
采用shannon′s多样性指数(H′)评价罗田甜柿和野柿各表型性状在所选群体间的表型多样性,结果(表2)表明:罗田甜柿13个表型性状的shannon′s多样性指数变化范围为1.38~2.09,平均shannon′s多样性指数为1.80,其中,果实横径、种子宽、萼片大小等性状多样性指数较高,表明这些性状在每一级中分布较均匀,果实纵径、果形指数、种子质量等性状多样性指数较低,表明这些性状的表现型少,且在每个表现型上分布不均匀。野柿各表型性状在所选群体体间的shannon′s多样性指数(H′)变化范围为1.51~2.28,平均shannon′s多样性指数为2.04,其中,单果质量、果实横径、柿蒂大小、萼片大小、种子数、种子宽、种形指数、种子质量、可溶性固形物含量等多个性状多样性指数较高,表明这些性状蕴藏着丰富的变异,果形指数、种子长等性状多样性指数较低,说明这些性状在群体间较稳定。

表2 大别山区两种甜柿表型性状变异Table 2 Genetic variation of D.kaki and D.kaki var.silvestris
2.3 罗田甜柿和野柿不同群体种实表型性状差异
不同群体罗田甜柿种实性状多重比较结果(表3)显示:湖北罗田云架山(P3)群体果实的单果质量、果实纵径等指标高于其它群体,湖北麻城盐田河(P6)群体果实横径高于其余群体,但各群体间差异均未达到显著水平;
各群体果形指数为0.80~0.85,果实偏扁圆形;
各群体柿蒂大小差异不大,但萼片大小、种子数量及种子大小的差异较显著;
萼片最大的湖北麻城盐田河(P6)群体(20.32 mm)是最小的河南商城黄柏山(P7)群体(14.67 mm)的1.39倍,种子数量最多的湖北罗田平湖乡(P4)群体(5.03个)是最少的河南商城黄柏山(P7)群体(3.30个)的1.52倍;
各群体种子均为狭长形,种形指数为1.49~1.74,其中,河南商城黄柏山(P7)群体种子最长最宽,安徽金寨燕子河(P9)群体种子最短最窄,各群体种子质量的差异未达到显著水平;
各群体中湖北罗田丁家山(P5)群体果实硬度(7.86 kg·cm−2)高于其余群体,而安徽金寨燕子河(P9)群体的果实可溶性固形物含量(17.68%)高于其余群体。
不同群体野柿种实性状多重比较结果(表3)显示:各群体野柿果实以近圆形为主,少部分偏扁圆形,果形指数为0.80~1.03,果实大的似兵乓球大小,小的如桂圆大小,果实单果质量最大的湖北云架山(P3)群体(28.20 g)为最小的安徽霍山但家庙(P10)群体(13.27 g)的2.13倍,果实横径最大的湖北云架山(P3)群体(36.51 mm)为最小的安徽霍山但家庙(P10)群体(27.55 mm)的1.33倍,果实纵径最大的湖北麻城盐田河(P6)群体(37.78 mm)为最小的安徽霍山白马尖(P11)群体(27.57 mm)的1.37倍;
各群体中柿蒂最大的是湖北云架山(P3)群体(18.37 mm),最小的为河南商城黄柏山(P7)群体(13.89 mm),萼片最大的是湖北云架山(P3)群体(16.16 mm),最小的是安徽霍山白马尖(P11)群体(9.56 mm);
各群体种子数量存在差异,种子数量最多的为安徽霍山白马尖(P11)群体(6.20个),为最少的湖北罗田蕙兰山(P1)群体(2.18个)的2.84倍;
各群体种子均为狭长形,种形指数为1.70~2.06,各群体间种子长、种子宽、种形指数及种子质量均差异不显著,种子质量最大的湖北罗田蕙兰山(P1)群体(0.54 g)为最小的安徽金寨燕子河(P9)群体(0.34 g)的1.59倍;
各群体中,河南商城黄柏山(P7)群体的果实硬度(9.94 kg·cm-2)显著高于其余各群体,安徽霍山但家庙(P10)群体的果实可溶性固形物含量最高(26.59%),为最低的罗田蕙兰山(P1)群体(18.85%)的1.41倍。
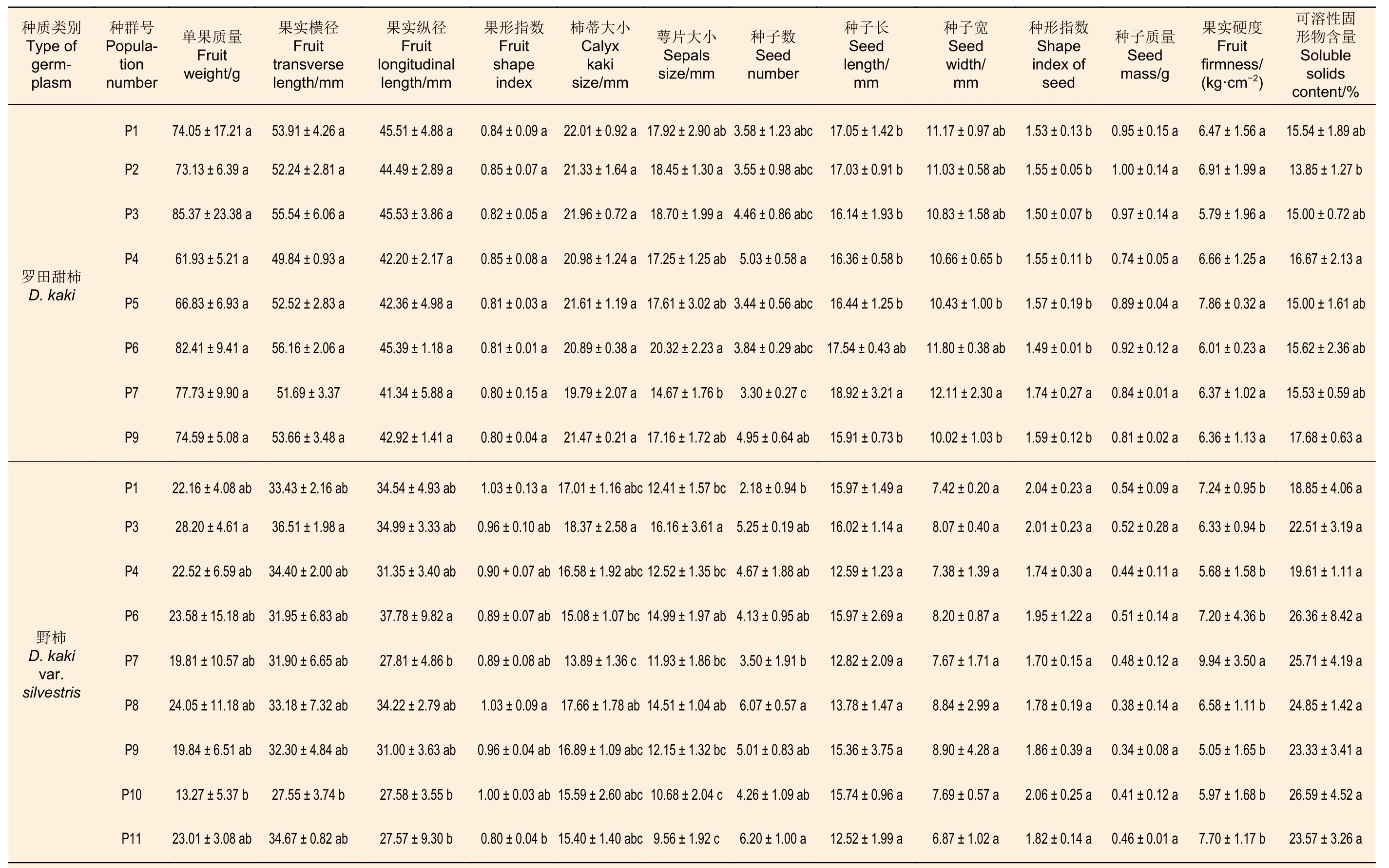
表3 罗田甜柿和野柿不同群体果实、种子性状变异Table 3 Phenotype characters of fruits and seeds in D.kaki and D.kaki var.silvestris populations
2.4 罗田甜柿和野柿种实表型性状分化差异
不同群体罗田甜柿和野柿的表型数据巢式方差分析结果(表4)显示:罗田甜柿群体间方差分量占总变异的17.07%,群体内方差分量占总变异的34.30%,说明群体间和群体内都发生一定的变异,群体内变异是变异的主要来源;
表型性状的平均表型分化系数为33.23%,表明群体间和群体内的变异贡献率分别为33.23%和66.77%,群体内的变异高于群体间的变异,其中果实纵径、果形指数、柿蒂大小、种子宽的表型分化系数较低,果实横径、萼片大小、种子数、种子长、可溶性固形物含量的表型分化系数较高。野柿群体间的方差分量占总变异的48.75%,群体内方差分量占总变异的6.69%,表明野柿群体的变异主要来源于群体间;
13个表型性状的表型分化系数为42.50%~平均表型分化系数为73.13%,表明野柿群体间和群体内的变异贡献率分别为73.13%和26.87%,群体间的表型有较大的分化,其中表型分化系数最大的为种子宽(96.22%)和果实硬度(90.73%),表型分化系数最小的为种子质量(42.50%)。

表4 罗田甜柿和野柿果实表型性状方差分量及群体间表型分化系数Table 4 Variance portions and differentation coefficients of fruit phenotype and quality traits among and within populations of D.kaki and D.kaki var.silvestris
罗田甜柿和野柿是大别山区2种主要的柿属植物资源,分布区存在一定的重叠,在形态上具有较大的差异。由于同域分布物种表型可塑性的式样和程度可能有很大的差异,它们通过表型分化减少种间竞争,进而使物种共存成为可能[22]。通常而言,局域分布的物种,其表型性状变异程度要远低于广泛分布的近缘种,如狭域种崂山腺齿越橘(Vaccinium oldhamii Miq.)[23]、新疆野生樱桃李(Prunus cerasifera Ehrh.)[24]等植物的果实变异程度显著低于其广布近缘种乌饭树(V.bracteatum Thumb.)[25]和野生樱桃李(P.divaricata LDB.)[26]。罗田甜柿各表型性状的平均变异系数为13.78%,处于较低变异水平。罗田甜柿是中国特有的甜柿品种,分布范围狭窄,由于人们对品质、产量等方面的要求,经过千百年来人工驯化改良,并通过嫁接等无性繁殖的方式将部分“优质基因”加以固定,导致含有其余性状基因的丢失和遗传基础的变窄,反应在表型性状上,就是表型多样性的降低和变异的减少[27];
而野柿的表型性状具有较高的变异,遗传基础较宽。这是由于野柿所处的生境相对复杂和多样,这就需要其具有更高的表型可塑性来提高对多变环境的响应能力。表型可塑性对于生物的分布有重要的意义,能使物种具有更宽的生态幅和更好的耐受性,可以占据更加广阔的地理范围和更加多样化的生境,成为生态位理论中的广幅种[28]。
表型性状中不同指标的可塑性具有极大的差异,与适合度相关性较强的一类性状(如种子形态等)可塑性一般较低,因为其受环境影响较小,相对较稳定。在对无患子(Sapindus spp.)[29]、国槐(Sophora japonica Linn.)[30]等植物的研究中,种子形态指数的平均变异系数分别为5.70%和6.27%,本研究中罗田甜柿和野柿种形指数的平均变异系数分别为8.35%和12.98%,仍处于较低的变异水平;
而其它间接影响适合度的性状,如种子质量等,其大小的变异是通过影响种子萌发进而影响物种的适合度,因此,具有相对较高的表型可塑性。罗田甜柿和野柿种子质量的平均变异系数分别为21.34%和30.94%,显著高于种子形态指标,与新疆野核桃(Juglans cathayensis Dode.)[31]、新疆野扁桃(Amygdalus ledebouriana Schleche.)[32]等植物的研究结果一致。
表型可塑性在植物对异质环境的适应过程中发挥了重要作用,它是植物适应异质生境的重要生态对策[33]。狭域种和广布种的进化适应性存在着差异。广布种由于生态幅宽,具有较高的种内多样性,表型可塑性强,因而对外界环境扰动的耐受性较强;
而狭域种由于其分布的特异性,生态幅较窄,形成了适应特定生境的表型,表型可塑性相对较低。大别山区罗田甜柿和野柿资源由于其进化适应性能力的差异,进而深刻影响着种群的扩散能力和种群动态,开展2种柿资源表型可塑性的比较研究,将有助于进一步理解广布种和狭域种的生活史对策。
作为大别山区广布种的野柿和狭域种的罗田甜柿,具有不同的表型可塑性。对果实和种子的表型性状进行比较分析表明,野柿的变异程度显著高于罗田甜柿。表现为野柿的表型性状的极差、变异系数、shannon′s多样性指数均显著高于罗田甜柿。罗田甜柿的变异主要来源于群体内,群体间分化较小;
而野柿群体间表型有较大的分化。表明这2种柿属植物资源具有不同的表型可塑性来适应不同的分布生境。






